Μερίδιο:



Ποιο είναι το πιρούνι αναπαραγωγής;
Το πιρούνι αναπαραγωγής είναι το σημείο στο οποίο λαμβάνει χώρα η αντιγραφή του DNA, ονομάζεται επίσης σημείο ανάπτυξης. Έχει ένα σχήμα Y, και καθώς προχωράει η αναπαραγωγή, η φουρκέτα μετατοπίζεται από το μόριο DNA.
Ο αναδιπλασιασμός του DNA είναι η κυτταρική διαδικασία που περιλαμβάνει την επανάληψη του γενετικού υλικού στο κύτταρο. Η δομή του DNA είναι μια διπλή έλικα και για να αναπαράγει το περιεχόμενό του πρέπει να ανοίξει. Κάθε ένας από τους κλώνους θα είναι μέρος της νέας αλυσίδας DNA, αφού η αναπαραγωγή είναι μια ημικονικοποιητική διαδικασία.
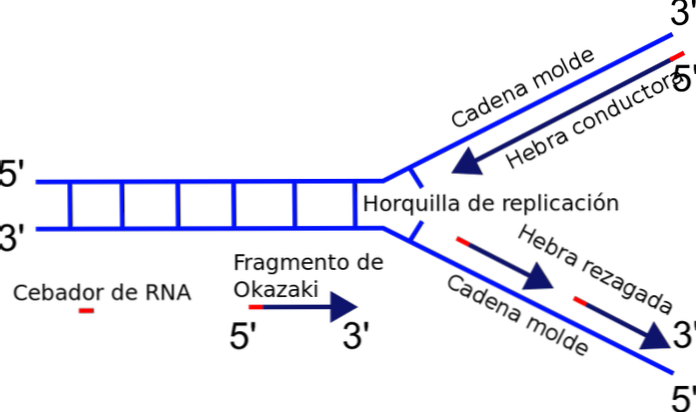
Το διχαλωτό διπλασιασμού σχηματίζεται ακριβώς μεταξύ της σύνδεσης μεταξύ των πρόσφατα διαχωρισμένων εκμαγείων προτύπου ή προτύπου και του διπλού DNA που δεν έχει ακόμη αντιγραφεί. Όταν αρχίζει η αντιγραφή του ϋΝΑ, ένας από τους κλώνους μπορεί εύκολα να αντιγραφεί, ενώ ο άλλος κλώνος αντιμετωπίζει ένα πρόβλημα πολικότητας.
Το ένζυμο που είναι υπεύθυνο για τον πολυμερισμό της αλυσίδας - η ϋΝΑ πολυμεράση - συνθέτει μόνο τον κλώνο του DNA στην κατεύθυνση 5'-3 '. Έτσι, ένας κλώνος είναι συνεχής και ο άλλος υφίσταται ασυνεχή ανατύπωση, δημιουργώντας θραύσματα του Okazaki.
Ευρετήριο
- 1 Ανατύπωση του DNA και του πιρουνιού αναπαραγωγής
- 1.1 Ανακατευθυνόμενη και αμφίδρομη αναπαραγωγή
- 1.2 Ένζυμα που εμπλέκονται
- 1.3 Έναρξη αναπαραγωγής και σχηματισμός περόνης
- 1.4 Επιμήκυνση και κίνηση του πιρουνιού
- 1.5 Τερματισμός
- 2 αντιγραφή DNA είναι ημι-συντηρητική
- 3 Το πρόβλημα πολικότητας
- 3.1 Πώς λειτουργεί η πολυμεράση?
- 3.2 Παραγωγή θραυσμάτων Okazaki
- 4 Αναφορές
Αναπαραγωγή του DNA και του πιρουνιού αναπαραγωγής
Το DNA είναι το μόριο που κατέχει την απαραίτητη γενετική πληροφορία όλων των ζωντανών οργανισμών - με εξαίρεση κάποιους ιούς.
Αυτό το μεγάλο πολυμερές που αποτελείται από τέσσερα διαφορετικά νουκλεοτίδια (Α, Τ, G και C) κατοικεί στον πυρήνα των ευκαρυωτικών, σε καθένα από τα κύτταρα που απαρτίζουν τους ιστούς αυτών των όντων (εκτός ώριμα ερυθρά αιμοσφαίρια των θηλαστικών, που στερούνται του πυρήνα).
Κάθε φορά που μια κυψέλη διαιρείται, το DNA πρέπει να αναδιπλασιαστεί προκειμένου να δημιουργηθεί ένα θυγατρικό κύτταρο με γενετικό υλικό.
Μονοκατευθυντική και αμφίδρομη αναπαραγωγή
Η αναπαραγωγή μπορεί να είναι μονόδρομη ή αμφίδρομη, ανάλογα με το σχηματισμό του πιρουνιού αναπαραγωγής στο σημείο προέλευσης.
Λογικά, στην περίπτωση της αντιγραφής σε μία κατεύθυνση, σχηματίζεται μόνο μία διχάλα, ενώ σε αμφίδρομη αναπαραγωγή σχηματίζονται δύο πιρούνια.
Συμπεριλαμβάνονται τα ένζυμα
Για αυτή τη διαδικασία απαιτείται ένα πολύπλοκο ενζυμικό μηχανισμό, το οποίο λειτουργεί γρήγορα και μπορεί να αναπαράγει το DNA με έναν ακριβή τρόπο. Τα σημαντικότερα ένζυμα είναι ϋΝΑ πολυμεράση, DNA πριμάση, ελικάση ϋΝΑ, ϋΝΑ λιγάση και τοποϊσομεράση.
Έναρξη της αναπαραγωγής και σχηματισμός του πιρουνιού
Ο αναδιπλασιασμός του DNA δεν ξεκινά από οποιαδήποτε τυχαία θέση στο μόριο. Υπάρχουν συγκεκριμένες περιοχές στο DNA που σηματοδοτούν την αρχή της αντιγραφής.
Στα περισσότερα βακτήρια, το βακτηριακό χρωμόσωμα έχει ένα μοναδικό σημείο εκκίνησης πλούσιο σε ΑΤ. Αυτή η σύνθεση είναι λογική, αφού διευκολύνει το άνοιγμα της περιοχής (τα ζεύγη AT συνδυάζονται με δύο γέφυρες υδρογόνου, ενώ το ζεύγος GC με τρία).
Καθώς το DNA αρχίζει να ανοίγει, σχηματίζεται μια δομή σχήματος Υ: το πιρούνι του αναδιπλασιασμού.
Επιμήκυνση και κίνηση του πιρουνιού
Η ϋΝΑ πολυμεράση δεν μπορεί να ξεκινήσει τη σύνθεση των θυγατρικών αλυσίδων από την αρχή. Χρειάζεστε ένα μόριο που έχει 3'-άκρο έτσι ώστε η πολυμεράση να έχει το σημείο να αρχίσει να πολυμερίζεται.
Αυτό το ελεύθερο 3 'άκρο προσφέρεται από ένα μικρό μόριο νουκλεοτιδίων που ονομάζεται εκκινητής ή εκκινητής. Το πρώτο ενεργεί ως ένα είδος γάντζου για την πολυμεράση.
Με την πορεία αναπαραγωγής, το πιρούνι αναπαραγωγής έχει τη δυνατότητα να μετακινηθεί κατά μήκος του DNA. Το βήμα της διχαλωτής διχοτόμησης αφήνει δύο μόρια ϋΝΑ μονής ζώνης που κατευθύνουν το σχηματισμό των θυγατρικών μορίων διπλής ζώνης.
Το πιρούνι μπορεί να προωθηθεί χάρη στη δράση των ενζύμων ελικάσης που εκτοπίζουν το μόριο DNA. Αυτό το ένζυμο θραύει τους δεσμούς υδρογόνου μεταξύ των ζευγών βάσης και επιτρέπει την μετατόπιση του περονιού.
Τερματισμός
Η αναπαραγωγή τερματίζεται όταν οι δύο πιρούνες βρίσκονται στους 180 ° C από την προέλευση.
Σε αυτήν την περίπτωση, μιλάμε για όσο ρέει η διαδικασία της αντιγραφής σε βακτηρίδια και θα πρέπει να σημειωθεί τη διαδικασία της συστροφής της κυκλικό μόριο που περιλαμβάνει την αντιγραφή. Οι τοποϊσομεράσες παίζουν σημαντικό ρόλο στην αποδέσμευση του μορίου.
Ο αναδιπλασιασμός του DNA είναι ημι-συντηρητικός
Έχετε αναρωτηθεί ποτέ πώς γίνεται η αναπαραγωγή στο DNA; Δηλαδή, μια άλλη διπλή έλικα πρέπει να προκύψει από τη διπλή έλικα, αλλά πώς συμβαίνει αυτό; Για αρκετά χρόνια, αυτό ήταν ένα ανοιχτό ερώτημα μεταξύ των βιολόγων. Θα μπορούσαν να υπάρξουν πολλές μεταβολές: δύο παλιές δέσμες μαζί και δύο νέες μαζί, ή ένα νέο νήμα και ένα παλιό για να σχηματίσουν τη διπλή έλικα.
Το 1957, η ερώτηση αυτή απαντήθηκε από τους ερευνητές Matthew Meselson και Franklin Stahl. Το μοντέλο αναπαραγωγής που προτάθηκε από τους συγγραφείς ήταν το ημικυκνωτικό.
Οι Meselson και Stahl δήλωσαν ότι το αποτέλεσμα της αντιγραφής είναι δύο δίκλωνες ϋΝΑ μόρια. Κάθε ένα από τα προκύπτοντα μόρια αποτελείται από έναν παλαιό κλώνο (από τη μητέρα ή το αρχικό μόριο) και από ένα νέο συνθετικό νέο κλώνο..
Το πρόβλημα της πολικότητας
Πώς λειτουργεί η πολυμεράση?
Η έλικα ϋΝΑ σχηματίζεται από δύο αλυσίδες που εκτελούνται με αντιπαράλληλο τρόπο: ο ένας πηγαίνει στην κατεύθυνση 5'-3 'και ένας άλλος 3'-5'.
Το πιο σημαντικό ένζυμο στη διαδικασία αντιγραφής είναι η DNA πολυμεράση, η οποία είναι υπεύθυνη για την καταλυτική σύνδεση των νέων νουκλεοτιδίων που θα προστεθούν στην αλυσίδα. Η πολυμεράση DNA μπορεί να επεκτείνει μόνο την αλυσίδα στην κατεύθυνση 5'-3 '. Αυτό το γεγονός εμποδίζει την ταυτόχρονη αλληλεπικάλυψη των αλυσίδων στο πιρούνι αναπαραγωγής.
Γιατί; Η προσθήκη νουκλεοτιδίων συμβαίνει στο ελεύθερο άκρο 3 'όπου βρίσκεται μία ομάδα υδροξυλίου (-ΟΗ). Έτσι, μόνο μία από τις αλυσίδες μπορεί να ενισχυθεί εύκολα με την τελική προσθήκη του νουκλεοτιδίου στο 3 'άκρο. Αυτό ονομάζεται αγώγιμο ή συνεχές σκέλος.
Παραγωγή θραυσμάτων Okazaki
Ο άλλος κλώνος δεν μπορεί να επιμηκυνθεί, επειδή το ελεύθερο άκρο είναι 5 'και όχι 3' και καμία πολυμεράση δεν καταλύει την προσθήκη νουκλεοτιδίων στο άκρο 5 '. Το πρόβλημα επιλύεται με τη σύνθεση πολλαπλών βραχέων θραυσμάτων (130 έως 200 νουκλεοτιδίων), καθένα στην κανονική διεύθυνση αντιγραφής από 5 'έως 3'.
Αυτή η ασυνεχής σύνθεση θραυσμάτων τελειώνει με την ένωση καθενός από τα μέρη, μια αντίδραση που καταλύεται από λιγάση ϋΝΑ. Προς τιμήν του ανακαλύπτρου αυτού του μηχανισμού, του Reiji Okazaki, τα μικρά τμήματα που συντίθενται καλούνται θραύσματα του Okazaki.
Αναφορές
- Alberts, Β., Bray, D., Hopkin, Κ., Johnson, A.D., Lewis, J., Raff, Μ., & Walter, Ρ. (2015). Βασική βιολογία των κυττάρων. Garland Science.
- Cann, Ι.Κ., & Ishino, Υ. (1999). Αρχική αναπαραγωγή DNA: ταυτοποίηση των τεμαχίων για την επίλυση ενός παζλ. Γενετική, 152(4), 1249-67.
- Cooper, G. Μ., & Hausman, R. Ε. (2004). Το κελί: Μοριακή προσέγγιση. Medicinska naklada.
- Garcia-Diaz, Μ., & Bebenek, Κ. (2007). Πολλαπλές λειτουργίες πολυμεράσης ϋΝΑ. Κρίσιμες ανασκοπήσεις στις φυτικές επιστήμες, 26(2), 105-122.
- Lewin, Β. (2008). γονίδια IX. Mc Graw-Hill Interamericana.
- Shcherbakova, Ρ. Ν., Bebenek, Κ., & Kunkel, Τ. Α. (2003). Λειτουργίες ευκαρυωτικών ϋΝΑ πολυμεράσεων. SAGE KE της επιστήμης, 2003(8), 3.
- Steitz, Τ. Α. (1999). DNA πολυμεράσες: δομική ποικιλομορφία και κοινοί μηχανισμοί. Journal of Biological Chemistry, 274(25), 17395-17398.
- Watson, J.D. (2006). Μοριακή βιολογία του γονιδίου. Ed. Panamericana Medical.
- Wu, S., Beard, W. Α., Pedersen, L. G., & Wilson, S. Η. (2013). Η δομική σύγκριση της αρχιτεκτονικής ϋΝΑ πολυμεράσης υποδηλώνει μια πύλη νουκλεοτιδίων προς την δραστική θέση της πολυμεράσης. Χημικές αναθεωρήσεις, 114(5), 2759-74.